CARACTERÍSTICAS FÍSICAS
Plumaje
Desempeñando un papel primordial a la hora de reconocer a cualesquier especie, y presentando a menudo adaptaciones especiales de uso al vuelo, el plumaje de un ave constituye el elemento de mayor importancia en la apariencia exterior de esta. De los poco más o menos 6 tipos existentes de plumas, las principales son las plumas de contorno y los plumones. Las remeras, timoneras, cubiertas del oído, y cubiertas de la cola son todas plumas de contorno, las cuales constituyen el plumaje visible del ave. Los plumones, por su parte, no son por lo general visibles, formando en la gran mayoría de las aves un plumaje oculto. Además de estos dos tipos, los loros (al igual que algunas otras aves) presentan un tipo particular de plumas, denominadas “plumones de polvo (o talco)”. Estas crecen continuamente, siendo plumones modificados cuyas barbas se desintegran formando un fino polvo, el cual le da al plumaje una apariencia muy propia. El polvo de los plumones sirve a las aves para limpiar sus plumas.
Las plumas de contorno nacen —en la gran mayoría de las aves— solamente de áreas determinadas en la piel del ave, llamadas pterilas. Las áreas interyacentes (apterias) se encuentran ya sea desnudas o cubiertas con plumones, aunque ocultas por las plumas de contorno. Existe una distribución definida de las pterilas (excepto en los pingüinos, tucanes, y aves corredoras). A todo esto, la pterilografía (útil en ocasiones dentro de la taxonomía) es conocida como la ciencia que se encarga del estudio de los patrones de nacimiento de las plumas en las aves.
Las pterilas en los loros se encuentran ralamente distribuidas, y con ello las áreas ápteras son extensas. Con excepción del pescuezo, en donde en muchas especies las apterias se encuentran desnudas, de toda la piel del ave nacen plumones en profusión.
Color
Existen dos tipos de colores en las aves: colores estructurales, cuyas estructuras responsables se encuentran presentes en las barbas y bárbulas de las plumas, y colores pigmentarios. Después de llevar a cabo estudios sobre el plumaje del Inseparable Cara de Durazno (Agapornis roseicollis), Dyck (1971) menciona que es a la reflexión de luz de los cilindros de queratina, los cuales constituyen la estructura esponjosa de las barbas, a lo que se deben los colores azules y verdiazules en las plumas de esta especie y, tal vez, de la mayoría de las demás especies. Al variar el tamaño de la estructura esponjosa, es posible que se produzcan colores diferentes en las barbas —la coloración producida por aquella puede no estar limitada a tonos azules y verdiazules. De acuerdo a Dyck, existe una diferencia entre las barbas verdes y las de color azul, la cual consiste en la presencia en las primeras de una corteza de pigmentación amarilla, así como de una estructura esponjosa más densa, con cilindros de queratina más anchos y menos aire en sus conductos. La torsión en ángulos rectos y el aplanamiento en porciones de las bárbulas producen, dice Vevers (1964), colores iridiscentes. Con respecto a los colores pigmentarios, estos se deben, haciendo su nombre obvio, a pigmentos. De estos, el más común es la melanina, cuya composición química no se conoce al 100%. La melanina puede ser, además de negra, café, siena, e incluso amarilla.
Dos tipos de pigmentos de composición desconocida se presentan en los loros. En uno de ellos, el pigmento es de color amarillo pálido al ser observado bajo la luz común, aunque presentándose dorado, color azufre, o verde bajo luz ultravioleta. Según Volker (1937), estos pigmentos se presentan comúnmente en especies pertenecientes a la región australoasiática, y en sólo dos especies del sur de América (Bolborhynchus lineola y Pionites leucogaster), no habiendo sido encontrados en la familia Loriidae ni en el género Eclectus. Algunas especies del género Cacatua presentan crestas coloreadas por uno de estos pigmentos. Pigmentos amarillos o rojos —no fluorescentes— constituyen el segundo tipo. Dos de las varias especies que presentan estos pigmentos son Melopsittacus undulatus, o Periquito Común, y Amazona leucocephala, o Loro Cubano.
La combinación de dos o más colores pigmentarios, así como de colores pigmentarios y estructurales, resulta en la presencia de muchos de los colores encontrados en los psitaciformes. La Cotorra de Cabeza Azul (Psittacula cyanocephala), señala Vevers, debe la coloración purpúrea de su cabeza al pigmento rojo contenido en las bárbulas de las plumas y a la estructura azul de las barbas.

Pico y lengua
Como ya se ha mencionado, el pico de un loro, por demás característico, se compone de dos mandíbulas, una superior y combada hacia abajo posada sobre una amplia inferior, combada esta hacia arriba. Ambas gozan de un amplio movimiento debido a la unión de la mandíbula superior al cráneo por medio de una estructura en forma de gozne o bisagra. Al movimiento de la mandíbula superior con respecto al cráneo se le llama cinesis; esta, aunque presente en la mayoría de las aves, es notable en los loros, los cuales se ven de esta manera habilitados para romper la cáscara de las semillas y frutas (como la nuez) de las que muchas especies se alimentan. La estructura básica del pico es idéntica en todas las especies, no obstante que existen ciertas modificaciones menores de acuerdo a los diferentes hábitos alimenticios. La especie Cacatua tenuirostris, o Corela de Pico Delgado, por ejemplo, posee un pico cuya mandíbula superior es alargada y menos curva, esto sirviendo a los requerimientos alimenticios de la especie (raíces y bulbos). De igual forma, las especies Enicognathus leptorhynchus y Purpureicephalus spurius (Conuro de Pico Delgado y Loro de Gorro Rojo) presentan una mandíbula superior alargada y menos curva, con la cual se ayudan en la extracción de ciertas semillas. Representantes dentro de la familia Loriidae, así como de los géneros Loriculus, Touit, y Brotogeris, los cuales se alimentan de polen y néctar, poseen picos sobresalientes y estrechos.
La lengua en los loros es gruesa y carnosa; esta, por lo general, presenta un epitelio grueso y calloso hacia la punta. Las papilas de la punta en las especies pertenecientes a la familia Loriidae, así como en la especie Lathamus discolor, utilizadas para recolectar polen, se presentan en forma de cerdas.
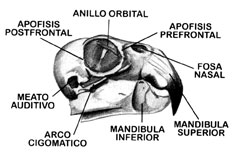
Cráneo
Amplio y relativamente grande, el cráneo de un loro presenta una espaciosa cavidad cerebral. Como regla, el cerebro en especies pequeñas es relativamente grande, y relativamente pequeño en especies grandes (Vlasblom, 1953). Las órbitas, bastante grandes, pueden presentar anillos óseos completos o incompletos, lo cual fue estudiado por Thompson (1900).
Sistema digestivo
Esencialmente un conducto en espiral, el cual va de la boca al ano, el sistema digestivo de un ave no es en base diferente al de todos los demás vertebrados. Los loros, al igual que otras aves consumidoras de granos, poseen un buche y una molleja bien desarrollados. El buche —en donde el alimento es almacenado para su posterior digestión, o para alimentar a las crías del ave por regurgitación— forma parte del esófago, siendo un área distensible y de paredes delgadas. En los mamíferos, el proventrículo y el ventrículo (o molleja) en conjunto conforman el estómago. A diferencia de otras especies, en las cuales la molleja se encuentra sumamente desarrollada, presentando paredes gruesas y músculos sólidos, los representantes de la familia Loriidae —y en especial aquellos que se alimentan casi por completo de néctar y polen— presentan una molleja débil y sin músculo. Las paredes glandulares del proventrículo son las encargadas de la secreción de jugos digestivos.
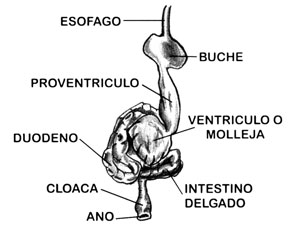
El Periquito Común (Melopsittacus undulatus) —se ha descubierto recientemente— secreta en su organismo una proteína similar a la producida por las células que revisten el buche en las palomas, y conocida como “leche de paloma”. En el Periquito Común, no obstante, dicha secreción puede originarse en el proventrículo.
HISTORIA NATURAL
En estado salvaje, los loros son por lo general difíciles de observar, pues habitan las cúpulas arbóreas y, como hemos visto, su color predominante es el verde. Una estridente bandada es a menudo lo único que se puede esperar al encontrarnos observando a estas aves en su ambiente natural. Sus colores cumplen su función, pasando desapercibidos durante las horas de comida, siendo hasta que rompen en repentino y estridente vuelo que el observador advierte su presencia. Existe, obviamente, más información acerca de los hábitos de especies las cuales se encuentran en abundancia en ciertos hábitats, o habitan en campo abierto. Las especies Platycercus eximius (Rosela Oriental), Psittacula eupatria (Cotorra Alejandrina), Poicephalus meyeri (Loro de Meyer), y Myiopsitta monachus (Cotorra Monje), por ejemplo, son bien conocidas, contándose con una buena cantidad de información sobre sus hábitos; en contraste, las costumbres de muchas de las aves habitantes de las selvas —como el Loro de los Higos de Salvadori (Psittaculirostris salvadorii), el Conuro de Plumas Doradas (Leptosittaca branickii), el Loro de Alas Manchadas (Touit stictoptera), y el Inseparable de Cuello Negro (Agapornis swinderniana)— son en sí desconocidas.
La información disponible con respecto a los hábitos o costumbres de crianza en los loros, además de generalizada, es en realidad pobre, al contrario de lo que sucede con casi todos los demás grupos de aves. Esto se debe a que la gran mayoría de los loros anida en sitios de difícil acceso (huecos en los árboles, orificios en termiteros) para la observación de los procesos de incubación, atención de los padres, desarrollo de los polluelos, etc. Las condiciones no naturales bajo las cuales son mantenidos los individuos en cautiverio pueden influir sobre las notables variaciones de comportamiento presentadas por estos, de cuya observación se deriva el conocimiento sobre la crianza en la mayoría de las especies.
LONGEVIDAD
En cautiverio, el promedio de vida en los loros es un asunto que se conoce bien. Este va de los 30 a los 50 años en especies grandes, habiéndose incluso registrado edades de más de 80 años. Forshaw (1973) habla de una Corelita (Cacatua sanguinea) la cual fue recolectada desde el nido en Australia central en 1904, encontrándose aún con vida y en cautiverio a la fecha de su mención. Australia es el único país en donde algunas especies han sido marcadas para su identificación, y por ello existe muy poca información con respecto a la longevidad de los loros en estado salvaje. Con base a estos datos se puede afirmar que los representantes del orden Psittaciformes —y en especial las especies más grandes— viven en promedio muchos años.
Al parecer, y como en el caso de muchas otras especies de aves, existe un más alto índice de mortalidad en loros jóvenes antes de alcanzar estos la madurez sexual. Las actividades humanas, así como el hombre mismo, son en la actualidad la principal causa de muerte en los loros; no es en absoluto inusual encontrarnos con individuos muertos a causa de disparos, trampas, autos, depredadores introducidos por el hombre, y tráfico de espacies. Las enfermedades, las aves de rapiña, y el hambre debida en ocasiones a condiciones climáticas desfavorables —como la sequía— son todas ellas causas naturales de muerte entre los psitaciformes.
En 1939 se observaron brotes de ornitosis fatal entre algunas especies de loros salvajes —Roselas Orientales (Platycercus eximus), en Tasmania, y Loros Rey Australianos (Alisterus scapularis), en Victoria; del mismo modo, una mortal epizootia se dio en la porción sudoriental de Australia del Sur entre una especie de loros no muy bien especificada, aunque podría haberse tratado del Loro de Rabadilla Roja (Psephotus haematonotus) (Burnet). De acuerdo a Miles (1959), la mortalidad de los loros (Platycercus adelaidae) que en enero de 1939 ocurrió en las proximidades de Adelaide, Australia del Sur, y la cual en aquel entonces fue atribuida a una extremadamente alta temperatura en la región, pudo haber formado parte de la ola epizoótica ocurrida durante aquel año. En 1887 y 1888 se dio una condición entre loros de la especie Psephotus haematonotus la cual impedía que las plumas fueran renovadas después de ser mudadas; los individuos afectados eran vistos correteando desnudos de un lado a otro en los suelos de Adelaide Hills, Australia del Sur, imposibilitados para volar y siendo presa fácil de los depredadores (Ashby, 1907). Más recientemente, y como producto de la enfermedad conocida como de Newcastle, la población de Conuros de Pico Delgado (Enicognathus leptorhynchus) en Chile se ha visto reducida; esta enfermedad ha afectado también a la Paloma Chilena (Columba araucana), aunque en este caso de manera más dramática, siendo la especie casi exterminada por completo por el virus (Johnson, 1967).
Las aves de rapiña atacan hasta a las especies grandes de loros. Forshaw (1973) reporta haber visto a una Cacatúa de Cresta Amarilla, o Cacatúa Galerita (Cacatua galerita) ser apresada por un Águila de Cola de Cuña (Aquila audax), aunque entonces pensando que aquella debía haberse encontrado débil y vieja; el mismo autor ha observado a Cacatúas Rosas, o Galahs (Eolophus roseicapillus) ser apresados por Halcones Peregrinos (Falco peregrinus). El Águila Pequeña (Hieraatus morphnoides), de hecho, ataca y mata a miembros de la especie E. roseicapillus.
DISTRIBUCIÓN
La gran mayoría de las especies pertenecientes al orden Psittaciformes habita en el hemisferio sur, principalmente en regiones tropicales. La especie que habita la máxima latitud norte es Psittacula himalayana, o Periquito del Himalaya, esto con la desaparición del Periquito de Carolina (Conuropsis carolinensis) en Estados Unidos. El Periquito del Himalaya es encontrado habitando el área de Safed Koh, en el este de Afganistán. El Conuro Austral, o Periquito de Magallanes (Enicognathus ferrugineus), por su parte, habita el extremo sur de la Tierra del Fuego, siendo el más sureño de los representantes del orden Psittaciformes a partir de la extinción del Periquito de Frente Roja (Cyanoramphus novaezelandiae) en la Isla Macquarie. Tanto la región australoasiática como Sudamérica son las áreas de mayor distribución en cuanto a loros se refiere. Existe en el sur de América, no obstante, una marcada uniformidad de tipos. Contrariamente a lo que se cree, la distribución de psitaciformes en Asia (subcontinente índico) y África es menor.
Distribución del orden Psittaciformes
La teoría del origen de las aves en la Antártida como un punto se ve respaldada, según Glenny (1954), por la actual distribución de los loros (las poblaciones australianas y sudamericanas, especialmente). Cracraft (1973), por su parte, y con base a la evolución presentada por las especies australoasiáticas y sudamericanas, apoya la teoría de que el origen de los psitaciformes se dio en el viejo mundo, y su posterior distribución ocurrió gracias a las interconexiones de los continentes del sur, esto hace de aproximadamente 70 a 90 millones de años, durante el cretáceo y el terciario temprano. Una mínima distribución de especies pudo haberse dado a través del Atlántico norte y el estrecho de Bering. Forshaw (1973) menciona que la posibilidad de que los loros se hayan originado en virtualmente toda región del globo no es en absoluto nula; la antigua proximidad de las tierras del sur entre sí representa sólo la existencia de rutas probables de dispersión, y no un centro de origen.
Mientras algunas especies de loros se encuentran confinadas a regiones específicas, la distribución de otras es bastante amplia. Este último es el caso de la Cotorra de Collar (Psittacula krameri), la especie con mayor distribución en el mundo. P. krameri habita en Asia y el norte de África, habiendo sido también introducida a regiones del Medio Este y el sudeste de Asia. El Loro de Cabeza Azul (Pionus menstruus), poblando de Centro a Sudamérica, es otra de estas especies, al igual que el Loro de Puerto Lincoln (Barnardius zonarius), habitante del oeste y sur de Australia. Las especies restringidas a zonas específicas habitan por lo general en pequeñas islas. Dos especies estrechamente relacionadas se encuentran habitando las Islas Antípodas, estas con un área total de aproximadamente 38 km²; de estas dos especies, el Periquito Verde de las Antípodas (Cyanoramphus unicolor) es una especie endémica. El Lori de Stephen (Vini stepheni), especie igualmente endémica, es encontrado poblando la Isla Henderson, un minúsculo atolón localizado en el Océano Pacífico. Aunque las especies encontradas sólo en pequeñas áreas dentro de grandes extensiones terrestres no son del todo comunes, sí existen algunas de ellas, como el Periquito de Frente Rufa (Bolborhynchus ferrugineifrons) y la especie Nannopsittaca panychlora.
HÁBITAT
Trepando por todos los árboles y acarreando frutos, los loros habitan principalmente en selvas tropicales bajas, en donde son particularmente numerosos. Forshaw (1973) observa que, en Nueva Guinea y el norte de Australia y Guyana, los loros concurren más comúnmente los límites de las selvas, en donde existen zonas delimitadas por ríos, pantanos, caminos, claros, etc., no poblando en grandes números las profundidades de la selva misma. Sin embargo, la dificultad para observarlos dentro de las selvas, agrega el autor, podría ser la razón de esta mayor observación en dichas zonas.
Una notable relación con los árboles presentan las especies encontradas en campo abierto, prefiriendo estas habitar zonas demarcadas por ríos, y siendo raramente vistas lejos de dichas zonas. Las incursiones de estas aves a jardines botánicos y parques son frecuentes; cabe señalar aquí que tales sitios se encuentran a menudo localizados dentro de límites urbanos. En algunas regiones de Australia es común observar a ciertas especies —Loro de Rabadilla Roja (Psephotus haematonotus) y Cacatúa Rosa o Galah (Eolophus roseicapillus), para ser precisos— posadas en líneas telegráficas en plena ciudad. En la India, pozos comunales localizados dentro de bulliciosos mercados sirven con frecuencia de bebedero a la Cotorra de Collar (Psittacula krameri). Forshaw (1973) menciona haber observado en São Paulo, Brasil, en un pequeño parque situado en la zona urbana y llamado Praça do República, pequeñas bandadas de Periquitos Llanos (Brotogeris tirica).
Los loros son encontrados, en general, menos comúnmente en altitudes mayores, presentándose —cuando lo hacen— en estos casos en contadas ocasiones en las zonas bajas. Algunas especies habitantes de las zonas altas son el Periquito Derbiana (Psittacula derbiana), del Tíbet, el Lori Papú (Charmosyna papou), de Nueva Guinea, el Loro de Cara Amarilla (Poicephalus flavifrons), de Etiopía, y el Periquito de la Sierra (Bolborhynchus aymara), de Sudamérica, entre otras, aunque de todos los representantes de las zonas altas, la especie más interesante es quizá el Kea (Nestor notabilis), de Nueva Zelanda. Esta especie vive en regiones montañosas (Alpes del Sur en Nueva Zelanda); se dice que llega a agredir y matar ovejas, aunque esto no ha sido comprobado. El Kea se pasea por la nieve en los alrededores de la zona, lo cual es en extremo inusual tomando en cuenta que los loros son un grupo mayormente tropical. Forshaw (1973), nuevamente, afirma haber observado en los Alpes del Sur en Australia a cacatúas Gang-Gang (Callocephalon fimbriatum) sobrevolar la cúpula arbórea durante nevadas, al igual que a roselas Carmesí (Platycercus elegans) buscando semillas al descubierto en superficies nevadas.
De las pocas especies confinadas a hábitats específicos, el Periquito Terrestre (Pezoporus wallicus) es una de las más notables. Esta especie, totalmente terrestre como su nombre lo indica, habita sólo en regiones cálidas de montaña y costeras y áreas inmediatas, al sur de Australia, un hábitat este bastante restringido, y el cual está rápidamente desapareciendo. |
